


Modification of adenosine to N6-methyladenosine (m6A) is a common post-transcriptional modification in most eukaryotic RNAs (including mRNA and miRNA); it regulates the stabilization of mRNA, its transport from the nucleus to the cytosol and its translational efficiency. Sperm formation is known to be associated with regulated changes in gene expression occurring before and during spermatogenesis. Several mechanisms operating in the testis to control gene expression are essential for the generation of sperm-specific transcripts and proteins. Recent information now documents key roles for m6A modification of RNA in testis. Gene knockout and mutations in methylases (writers) and demethylases (erasers) have been shown to result in male infertility. The intersection of two lines of investigation resulted in the discovery that FTO, one of two m6A erasers, appears to be regulated by the signaling enzyme GSK3α. Targeted disruption of GSK3α or its conditional knockout in developing spermatogenic cells results in male infertility. Epididymal maturation and the ability of sperm to fertilize eggs are impaired in knockout mice. Also, studies on signaling mechanisms underlying the maintenance of stem cell pluripotency identified a novel mechanism involving m6A demethylation by FTO and its regulation by GSK3. These two lines of evidence prompted preliminary studies which showed the presence of FTO, its binding to GSK3α, and reduction in the levels of m6A RNA in the testis of GSK3α knockout mice. Taken together these data form the basis for the hypothesis that regulation of FTO by GSK3α in developing spermatocytes and spermatids is essential for normal sperm function and male fertility.
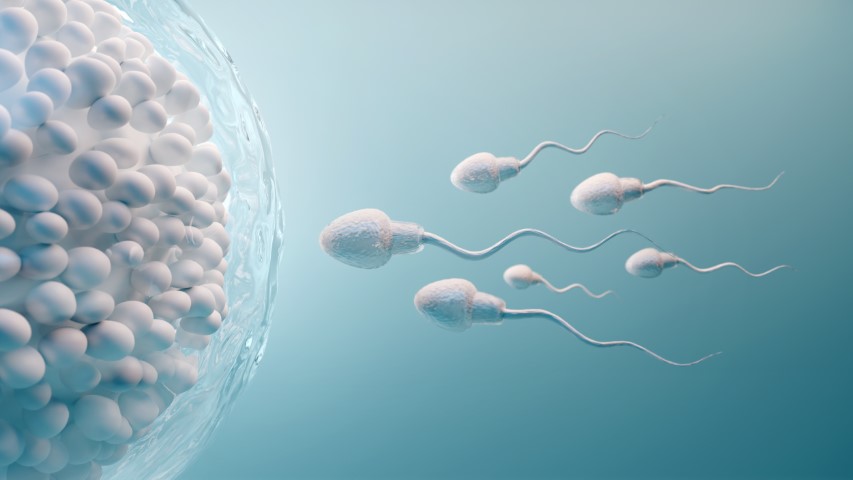
Deletion of testis/sperm-specific calcineurin isoform (viz. Ppp3r2/Ppp3cc) leads to male infertility in mice. Global or testis-specific disruption of the Gsk3α gene caused impaired sperm motility and infertility in the murine system. Inhibitors of calcineurin (viz. FK506 and cyclosporin A) and GSK3α have been shown to profoundly inhibit the ability of mouse sperm to fertilize eggs in vitro. FK506 injection into the mice (intraperitoneally) was found to induce reversible infertility in murine models. Therefore, the key questions now are whether similar inhibition of human seminal sperm calcineurin/GSK3α leads to a significant reduction in its fertilizing ability. Can this inhibition replicate the altered motility pattern, especially hyperactivation of human sperm cells as was observed in the case of mouse sperm? Can FK506 and/or BRD0705 be formulated into a potential reversible male contraceptive, in near future? We hypothesize that FK506 and BRD0705, both will be able to prevent sperm hyperactivation and thereby its ability to fertilize eggs.
The localization of GSK3α and GSK3β in mature sperm is different. Sperm, being a highly compartmentalized structure, doesn’t allow free movement of all its functional proteins across the entire cell. While GSK3β is only present in the anterior sperm head, GSK3α is chiefly localized in the sperm midpiece and tail, besides its minor presence in the anterior sperm head. This could be partly due to the dissimilar N- & C-terminal amino acid sequences of these paralogs, which may have contributed to their bimodal distribution subsequent to translation. Anyway, it results in differential substrate phosphorylation by these two kinase isoforms. Now, since deletion or inhibition of only GSK3α results in defective sperm motility (both progressive and during hyperactivation) and infertility, it is necessary to dissect its cognate substrates from a common pool of the same. We are employing multiple methods: the chemical genetics technique and the CRISPR-Cas9-dependent Gsk3 paralog-specific knockout mouse-based pharmacological inhibitor-based approach.
GSK3 is a worthwhile target to treat many cancer types. In childhood ALL, GSK3 has been implicated to have a major contribution. However, all the preclinical and clinical trials using GSK3 inhibitors in cancer that have been undertaken so far have shown side effects. It’s simply because of the lack of the paralog-selectivity of these GSK3 inhibitors; examples of these failed GSK3 blockers include Tideglusib, CHIR99021, 9-ING-41, LY2090314, etc. With the help of paralogue-selective GSK3 inhibitors, viz., BRD0705 against GSK3α & BRD3731 against GSK3β, we plan to identify the ‘hyperactive’ GSK3 isoform in different human cancer cell lines and clinical ALL samples and develop mice CDX/PDX models for anti-cancer drug screening.
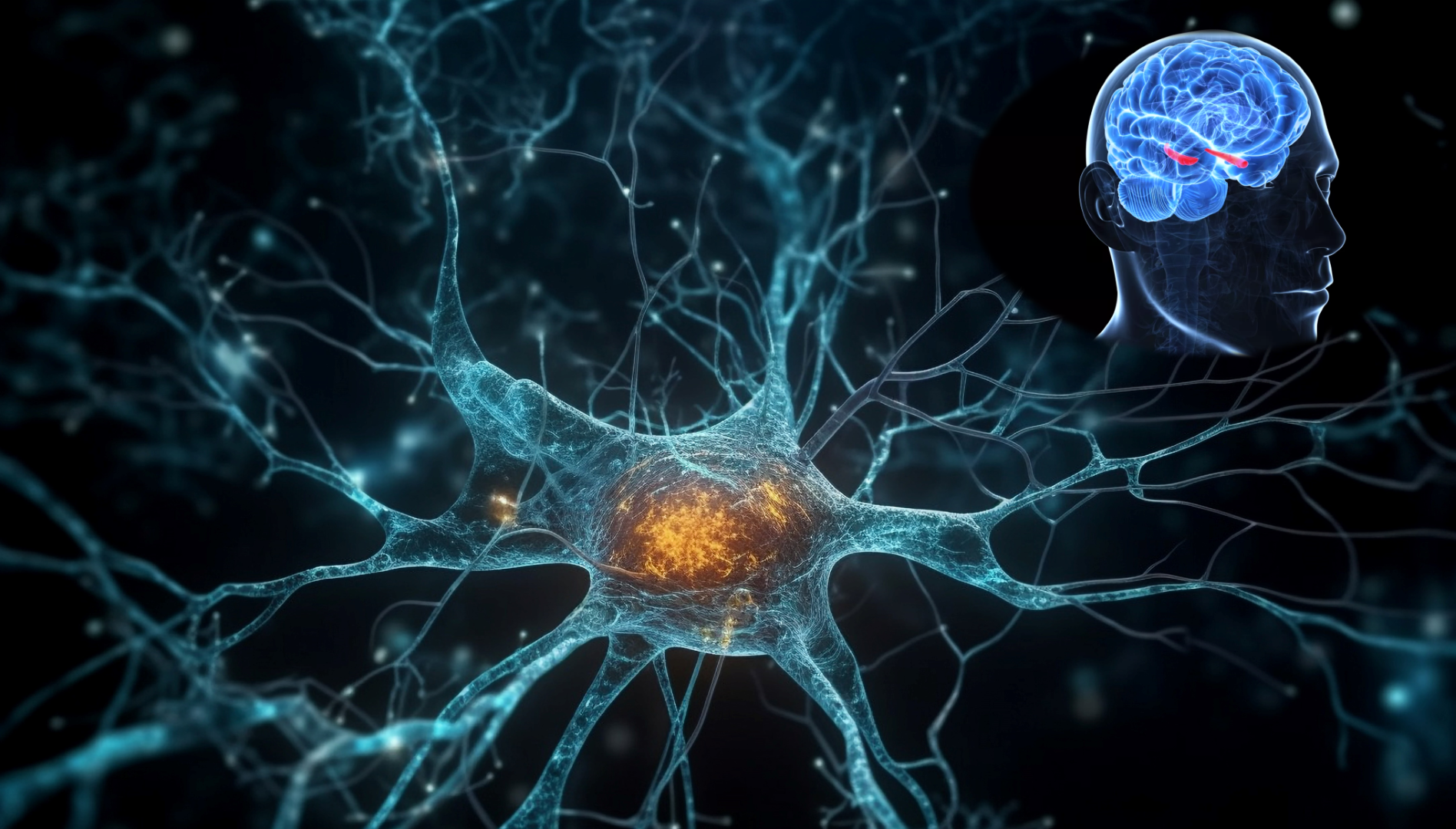
GSK3 plays a vital role in various neurodegenerative illnesses, particularly those that are primarily linked to aging, such as Alzheimer’s disease (AD) and Parkinson’s disease (PD). The dysregulation of GSK3 is responsible for many of the clinical symptoms of AD in both sporadic and familial cases. Therefore, GSK3 has been suggested as a potential target for the development of new AD treatments. Over-activation of GSK3 causes tau hyper-phosphorylation and increases amyloid beta production in a bidirectional manner. The involvement of a particular isoform of GSK3 in the pathophysiology of AD remains unknown. We propose selective targeting of overexpressed GSK3 paralogs can help in mitigating AD symptoms with minimal side effects.